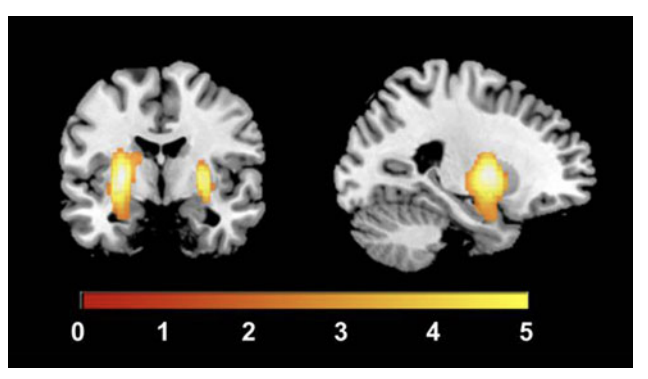
״We present evidence that support a model by which alterations in HRV might precede the onset of depression. This evidence suggests that the interplay between autonomic processes and alterations in emotional regulation can serve as a potential risk factor for depression.״
A highly replicable decline in mood during rest and simple tasks.
Jangraw D, Keren H, Sun H, Bedder RL, Rutledge RB, Pereira F, Thomas AG, Pine DS, Zheng C, Nielson DM, Stringaris A. Nature Human Behavior, 2023 link
״We show that accounting for time using a linear term significantly improves the fit of a computational model of mood. Our work provides conceptual and methodological reasons for researchers to account for time’s effects when studying mood and behavior.״
Emotion-related impulsivity and risky decision-making: A systematic review and meta-regression
Elliott MV, Johnson SL, Pearlstein JG, Muñoz Lopez DE, Keren H. Clinical Psychology Review, 2023 link
“the data suggest that risky decision-making is a cognitive domain that relates to emotion-related impulsivity. We conclude with recommendations regarding the specific types of tasks and arousal inductions that will best capture emotion-related impulsivity in future experimental research.”
Neural responses to reward valence and magnitude from pre-to early adolescence.
Gadassi Polack R, Mollick JA, Keren H, Joormann J, Watts R. NeuroImage, 2023 link
“Our results suggest sub-specialization to valence vs. magnitude within many ROIs of the reward circuitry. Additionally, in line with theoretical models of adolescent development, our results suggest that the ability to benefit from success increases from pre- to early adolescence.“
The temporal representation of experience in subjective mood reports.
Keren H, Zheng C, Jangraw D, Chang K, Vitale A, Nielson D, Rutledge R B., Pereira F, Stringaris A. eLife, 2021 link,Featured on the journal website
“We show that a Primacy model accounts better for mood reports compared to a range of alternative temporal representations across random, consistent, or dynamic reward environments, different age groups, and in both healthy and depressed participants.”
“…this is the first study to harness computational models of mood to relate mood fluctuations to variations in neural oscillations with MEG.”
Great Expectations: A Critical Review of and Suggestions for the Study of Reward Processing as a Cause and Predictor of Depression
Nielson DM, Keren H, O’Callaghan G, Jackson S, Douka I, Zheng CY, Vidal-Ribas P, Pornpattananangkul N, Camp C, Gorham L, Wei C, Kirwan S, Stringaris A. Biological Psychiatry, 2020 link “We found that reward processing abnormalities do not reach levels that would be useful for clinical prediction, yet the available evidence does not preclude a possible causal role in depression.”
Unstructured network topology begets order-based representation by privileged neurons
Bauermeister C, Keren H, Braun J
Biological Cybernetics, 2020 link “In conclusion, we offer a minimal model for the representational role of pioneer neurons, as observed experimentally in vitro. In addition, we show that broadly heterogeneous connectivity enhances the representational capacity of unstructured networks.”
Bidirectional associations between stress and reward processing in children and adolescents: a longitudinal neuroimaging study
Vidal-Ribas P, Benson B , Vitale AD, Keren H, Harrewijn A, Fox NA, Pine DS, Stringaris A.
Biological Psychiatry: Cognitive Neuroscience and Neuroimaging, 2019 link “Our findings provide support for bidirectional associations between stress and reward processing, in that stress may impact reward anticipation, but also in that reduced reward anticipation may increase susceptibility to stress.”
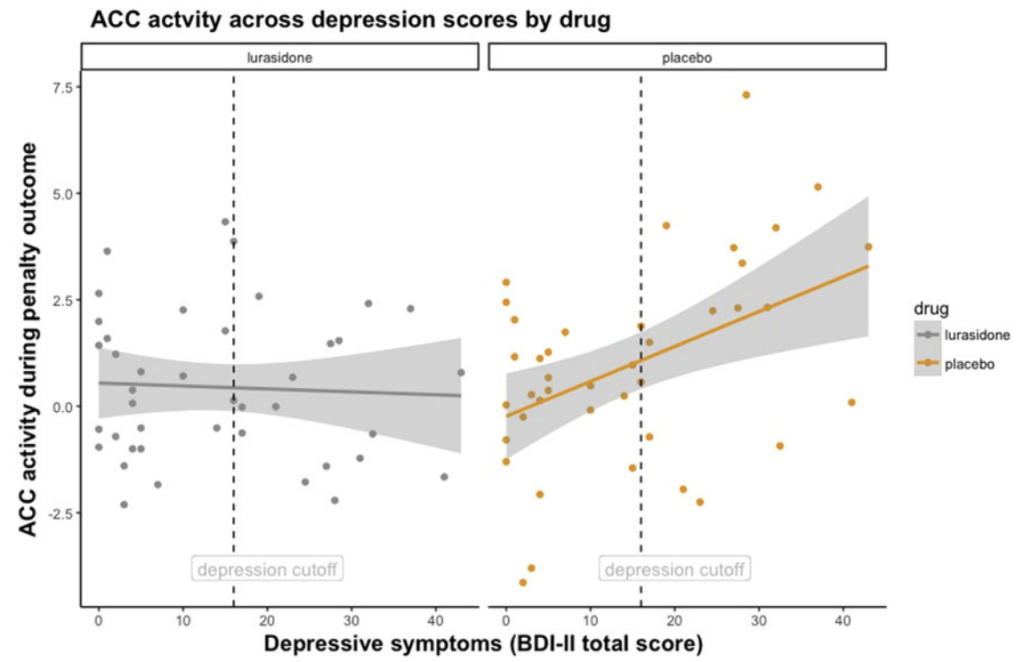
Modulation of anterior cingulate cortex reward and penalty signalling in medication-naive young-adult subjects with depressive symptoms following acute dose lurasidone
Wolke SA, Mehta MA, O’Daly O, Zelaya F, Zahreddine N, Keren H, O’Callaghan G, Young AH, Leibenluft E, Pine DS, Stringaris A.
Psychological Medicine, 2019 link “Lurasidone acutely normalises reward processing signals in individuals with depressive symptoms. Lurasidone’s antidepressant effects may arise from reducing responses to penalty outcomes in individuals with depressive symptoms.”
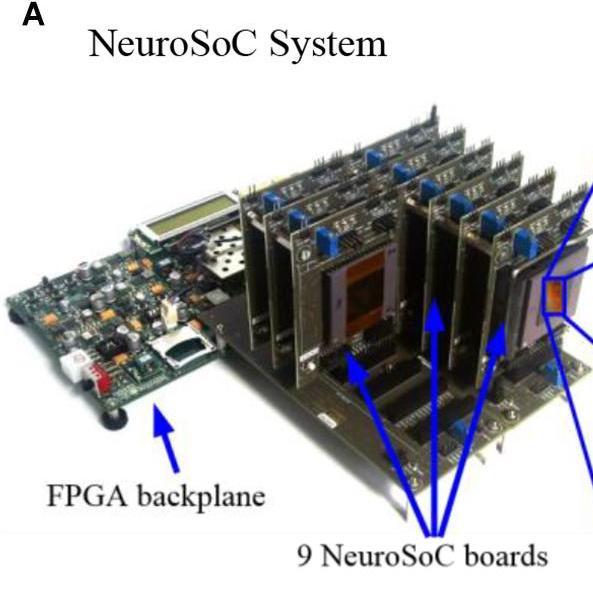
A biohybrid setup for coupling biological and neuromorphic neural networks.
Keren H, Partzsch J, Marom S, Mayr C. Frontiers in Neuroscience, 2019 link “we provide an experimental model for neuromorphic-neural interfaces, hopefully to advance the capability to interface with neural activity, and with its irregularities in pathology.”
Featured and republished in the Neural Technology Editors’ Pick, 2021,link to the article
Reward processing in depression: a conceptual and meta-analytic review across electrophysiological and fMRI studies
Keren H, O’Callaghan G, Vidal-Ribas P, Buzzell G, Brotman MA, Leibenluft E, Pan P, Meffert L, Kaiser A, Wolke S, Pine DS, Stringaris A.
American Journal of Psychiatry, 2018 link “..the findings show consistent neural aberrations during reward processing in depression, namely, reduced striatal signal during feedback and blunted FRN. These aberrations may underlie the pathogenesis of depression and have important implications for development of new treatments.”
Is the encoding of Reward Prediction Error reliable during development?
Keren H, Chen G, Benson B, Ernst M, Leibenluft E, Fox NA, Pine DS, Stringaris A.
NeuroImage, 2018 link “Positive RPEs are found to be encoded in the striatum and negative values in the insula. We show that the RPE signal is reliable mostly in the insula, across a few months and a few years. Our findings indicate that fMRI RPE signals can be used as biomarkers in longitudinal studies of health and disease.”
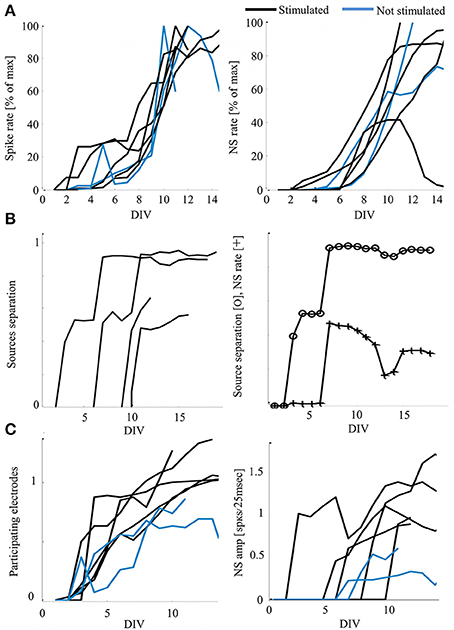
Development of Cortical Networks under Continuous Stimulation
Orenstein O, Keren H.
Front. Mol. Neurosci, 2017 link “This work, extends these studies in the direction of continuous recording and stimulation of cortical networks in the course of development, as of the second day in-vitro. The experimental database is comprised of six networks developing while exposed to continuous stimulation, as well as an additional two non-stimulated networks, similarly monitored.”
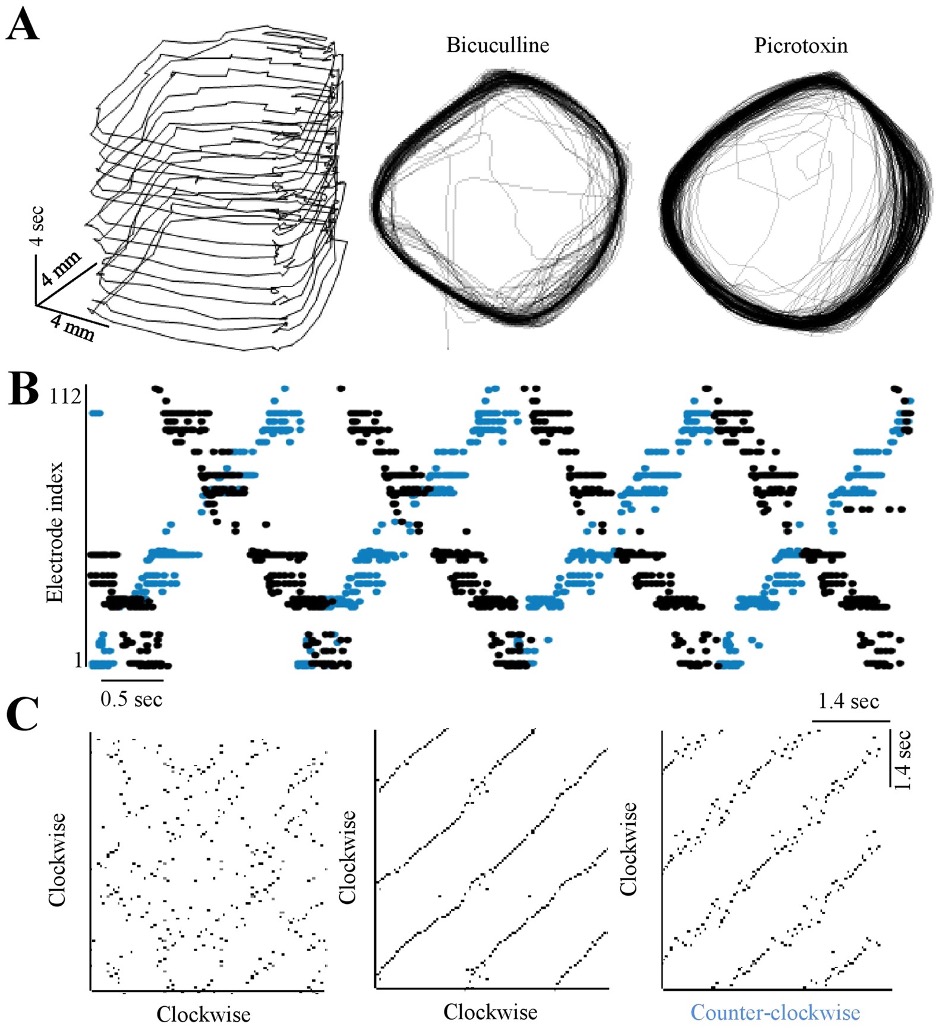
Long-range synchrony and emergence of neural reentry
Keren H, Marom S
Nature Scientific Reports, 2016 link “When λ is in the range of the longest propagation path in a random network, dictated by the network dimensions, the network tends to fall into a self-sustained reentrant mode of synchronous activity. We further show that the length scale is sensitive, in a non-trivial manner, to the level of inhibitory transmission.”
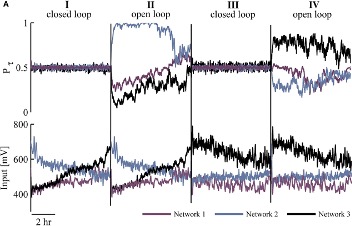
Controlling neural network responsiveness: tradeoffs and constraints
Keren H, Marom S
Frontiers in Neuroengineering, 2016 link “we show that enforcement of stable high activity rates by means of closed loop control may enhance alteration of underlying global input–output relations and activity dependent dispersion of neuronal pair-wise correlations across the network.”
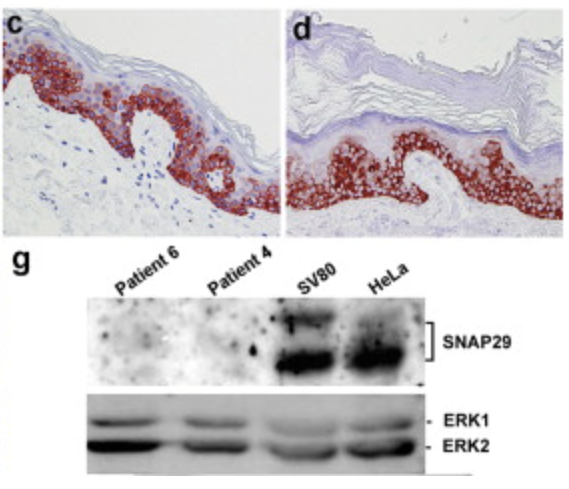
A Mutation in SNAP29, Coding for a SNARE Protein Involved in Intracellular Trafficking, Causes a Novel Neurocutaneous Syndrome Characterized by Cerebral Dysgenesis, Neuropathy, Ichthyosis, and Palmoplantar Keratoderma
Sprecher E, Ishida-Yamamoto A, Mizrahi-Koren M, Rapaport D, Goldsher D, Indelman M, Topaz O, Chefetz I, Keren H, O’Brien TJ, Bercovich D, Shalev S, Geiger D, Bergman R, Horowitz M, Mandel H. The American Journal of Human Genetics, 2005 link “These data underscore the importance of vesicle trafficking regulatory mechanisms for proper neuroectodermal differentiation.”
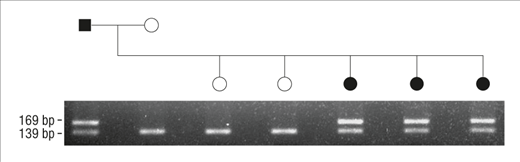
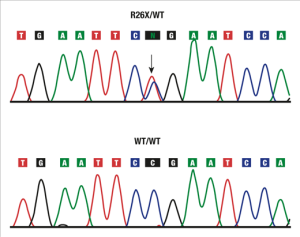
Diffuse Nonepidermolytic Palmoplantar Keratoderma Caused by a Recurrent Nonsense Mutation in DSG1
Keren H, Bergman R, Mizrahi-Koren M, Kashi Y, Sprecher E.
“Mutation analysis eventually led to the identification of a causative recurrent nonsense mutation in this gene. The present study illustrates the efficacy of an integrative diagnostic approach to palmoplantar keratodermas involving clinical assessment, pathologic examination, microsatellite marker screening, and mutational analysis.”

")